The subject was FOXP2 and the evolution of speech. FOXP2 is a gene that has been linked to some faculties of speech and language, the media going as far, as they do, as calling it "the speech gene". It isn't a terribly current or "right now" topic, but it highlights many aspects of evolutionary neuroscience and it spans everything from genetics and development to evolution to behavior and society so it lends itself to interesting discussion.
I wrote down some questions to use as a guidance in the discussion but I didn't hand them out to the students beforehand 'cause I thought that they'd be more free to take the discussion wherever they wanted, it being such a wide-spanning subject. Apparently that was a mistake. The first group of students was very quiet, frustratingly so, and I had to do most of the talking. I think they found the articles difficult. I can understand that, but they were supposed to be challenging. The purpose of the seminar is to give some training in critical reading of scientific literature. This does take some effort and maybe they would have needed some concise questions to guide them. I gave the second group the questions and 15 minutes to prepare some answers at the beginning of the seminar and they did so much better. So that's something to consider for the next time.
I centered the seminar around a general review article from a few years back, a couple of short reports on recent FOXP2 evolution, one of which is about the controversial subject of Neanderthal gene sequencing, and a really nice article from last year on FOXP2 in echolocating bats (first 4 references below). All the students got the review article and one half got the reports while the other half got the bat article. My thought was that we could take the discussion from the anthropocentric notion of FOXP2 having evolved in the human lineage to produce our "superior" faculties of speech and language and warp it towards bats and how FOXP2 seems to have evolved in their lineage as well to produce their unique use of vocalization for echolocation. Along the way we also discussed some general concepts of molecular evolution, the neuroanatomy of speech production, the definition of speech and language, the emergence of culture and the selective advantages of evolving such a complex means of communication.
First some introduction.
The FOXP2 gene was first identified through the study of the so called "KE family" in the late 90s. This is a family in which a severe speech and language disorder affects almost half of the members. This case was interesting because the disorder segregated from generation to generation in a pattern that pointed very decisively towards there being just one gene causing it. You can see a pedigree of the KE family below. Shaded symbols indicate affected individuals.
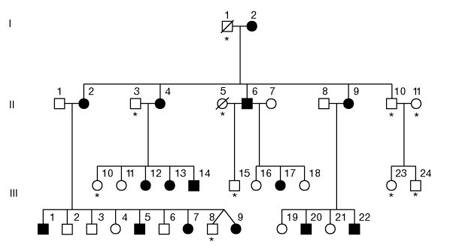
Ref: S.E. Fisher et. al (see reference below)
The researchers were able to identify the region on chromosome 7 that was associated with the speech disorder and subsequently they were able to identify the gene. Even though FOXP2 should not by any means be called "the speech gene", its identification and study has given us an "entry way" into the very complex processes that govern speech and ultimately perhaps language.
These are the questions I prepared for the seminar with my own answers, adapted for an undergraduate biology crowd.
1. What is the function of the FOXP2 gene product? How does it act?
FOXP2 is a transcription factor, a protein that binds to DNA and regulates the expression of a variety of specific genes. In the case of FOXP2 it's still unknown which ones. FOXP2 in particular represses the expression of genes. The fact that it's a transcription factor puts it in a place of particular interest for several reasons. (1) It makes it probable that it's central to the processes that underlie speech. (2) Small changes in a transcription factor can give rise to major innovations because they influence a wide variety of genes and thus functions. (3) Transcription factors have dual roles - they act during development, setting up structures and functions, as well as in the mature organism, regulating the same. This mirrors the complex development and plasticity of speech.
2. FOXP2 is an extremely conserved gene in vertebrates. What does this mean? Can you relate this fact to the FOXP2 protein function?
The word conserved means that there is very little sequence diversity between lineages. In other words, there is extremely little difference between the crocodile FOXP2, chicken FOXP2 and human FOXP2. Transcription factors are generally more conserved since they have a very basic function - even the smallest change could have enormous consequences since transcription factors regulate a wide variety of genes and functions, but also because they act during development.
3. FOXP2 was related to speech by the study of individuals with speech abnormalities (particularly the KE family). How are their FOXP2 genes aberrant?
The KE family has a substitution from arginine to histidine on position 553. This affects the DNA-binding region of the protein, leaving it useless. Another patient has a premature stop codon that leaves the protein too short to function and yet another patient has a translocation in the region containing the gene.
4. Why is it useful to study FOXP2 in vocal learning birds and bats? What are the main findings in this regard?
The implication of FOXP2 in brain regions involved in bird song learning are considered parallel to the human vocal learning of language. Young birds, just like us, mimic the sounds that adults make in order to learn them. It has been found that FOXP2 expression is increased in periods of song learning. These seasonal periods of plasticity could mirror the development of the neural circuits that make vocal learning possible. However no specific mutations have been related to this specific ability (including the study of vocal learning echolocating bats) and expression sites of FOXP2 are not different when comparing vocal learners with non-vocal learners.
5. In which brain regions is FOXP2 expressed? To what faculties of speech and language can you relate FOXP2 expression?
FOXP2 is expressed in similar and partly overlapping regions in all vertebrates studied, mainly in cerebellar and basal ganglia circuits. Regions include the lateral ganglionic eminence in the developing brain which gives rise to the striatal medium spiny projection neurons, involved in planning and modulation of movement; thalamus, in particular the regions that receive input from the basal ganglia; the inferior olives, which are part of the cerebellar motor learning and function; cerebellar Purkinje cells and deep cerebellar nuclei as well as sensory auditory midbrain structures. These regions implicate FOXP2 in the fine sensorimotor coordination/integration which underlies the sequenced behavior and learning behind speech and language.
FOXP2 is not expressed in the structures that form the trigeminal sensorimotor circuit that controls orofacial movements. So FOXP2 apparently has nothing to do with the control of the movements of the mouth and lower face, but rather with the coordination, planning and learning of these movements.
6. How is the human FOXP2 gene different from that of our closest extant relative the chimpanzee? What consequence does this have for the human FOXP2 protein function?
The human FOXP2 gene has two characteristic amino acid substitutions in exon 7 compared to the chimpanzee gene - a threonine to asparagine substitution in position 303 and an asparagine to serine substitution on position 325. The latter substitution leads to a hypothetical target site for protein kinase C and a minor structural change. Phosphorylation of transcription factors is an important way of regulating gene expression. Even this small difference between humans and chimpanzees could lead to a dramatic change in the regulations of the variety of genes that are under the control of FOXP2.
7. Has the human FOXP2 gene been under any selective pressure? How can we see this?
It seems as though the human FOXP2 gene has been under recent positive selection rather than relaxed negative selection. This means that whatever changes the human-specific mutations caused, they gave significant benefits to the individuals that carried them. This could be seen by studying the FOXP2 gene sequences in mouse, great apes and human and comparing non-synonymous substitutions, mutations that change the amino-acid sequence of the protein, with synonymous substitutions, mutations that don't.
8. The Neanderthal FOXP2 gene seems to be identical to that of modern humans. With other primates in mind, what consequences does this have for the expansion of modern humans?
This is a very big and very open discussion that in the end is more speculative than anything, albeit based on actual scientific findings. The question seems to be whether or not the emergence of a complex language lead to our cultural and therefore geographical and demographical expansion? It's definitely tempting to draw the conclusion that we became so successful because we evolved a language that allowed us to cooperate between individuals like never before. The human-specific mutations in FOXP2 may very well have been a pivotal point in this development. If Neanderthals had the same FOXP2 sequence as us modern humans, it's probable that they possessed many of the same faculties of language and culture that we do. Maybe at one time in history we understood each other?
9. In what way is FOXP2 special in echolocating bats? Can this provide any clues with regards to FOXP2 function? Can this be contrasted to FOXP2 in echolocating whales?
FOXP2 has diverged more in echolocating bats than any other group of vertebrates. This further implicates FOXP2 in sensorimotor coordination and vocal learning, which are requirements for echolocation. The same pattern could not be seen for echolocating whales, presumably because their echolocation is mediated through their foreheads and would not require sensorimotor coordination of their mouth and face.
10. Using what you have learned about FOXP2 in humans, birds, bats, whales and other animals, what can you say about the evolution of vocal communication and language? Is language a uniquely human feature?
This is yet another very big and very open discussion. Arguably, many animals possess some of the faculties of our speech and language. Most animals vocalize, for instance, even though it's only growls, barks, meows, shrieks, squeaks et.c. As we've seen, some learn their complex vocalization patters by mimicking adults, just like we do, and some even have more complex systems where different sounds are connected to different meanings. A few species of monkeys can distinguish between different predators and warn their fellow group-members accordingly.
But is this language then?
I would say no. The best definition I know of language is "the ability to produce infinite meaning from a finite sent of sounds or symbols", and this is clearly a human-specific feature, as far as our best knowledge goes. With our language we are able to describe not only that which we can sense, but also a wide spenctrum of things we cannot sense at all! Even things that are completely invented. We use language not only for utility but also creatively to make things up. We can even make up words that have no meaning attached to them.
Still, the fact that other animals possess some faculties of speech is significant because it firmly bases our unique adaptation of language and culture within natural processes by showing that a stage was already set from which a few genomic events could lead to our advanced use of vocalization.
Swedish blog tags: Vetenskap, Biologi, Neurovetenskap, Språk
Technorati tags: Science, Biology, Neuroscience, Language, FOXP2
Scharff, C., Haesler, S. (2005). An evolutionary perspective on FoxP2: strictly for the birds?. Current Opinion in Neurobiology, 15(6), 694-703. DOI: 10.1016/j.conb.2005.10.004
Enard, W., Przeworski, M., Fisher, S.E., Lai, C.S., Wiebe, V., Kitano, T., Monaco, A.P., Pääbo, S. (2002). Molecular evolution of FOXP2, a gene involved in speech and language. Nature, 418(6900), 869-872. DOI: 10.1038/nature01025
Krause, J., Lalueza-Fox, C., Orlando, L., Enard, W., Green, R., Burbano, H., Hublin, J., Hänni, C., Fortea, J., de la Rasilla, M., Bertranpetit, J., Rosas, A., Pääbo, S. (2007). The Derived FOXP2 Variant of Modern Humans Was Shared with Neandertals. Current Biology, 17(21), 1908-1912. DOI: 10.1016/j.cub.2007.10.008
Li, G., Wang, J., Rossiter, S.J., Jones, G., Zhang, S. (2007). Accelerated FoxP2 Evolution in Echolocating Bats. PLoS ONE, 2(9), e900. DOI: 10.1371/journal.pone.0000900
Fisher, S.E., Vargha-Khadem, F., Watkins, K.E., Monaco, A.P., Pembrey, M.E. (1998). Localisation of a gene implicated in a severe speech and language disorder. Nature Genetics, 18(2), 168-170. DOI: 10.1038/ng0298-168
Cecilia S. L. Lai, Simon E. Fisher, Jane A. Hurst, Faraneh Vargha-Khadem, Anthony P. Monaco (2001). A forkhead-domain gene is mutated in a severe speech and language disorder Nature, 413 (6855), 519-523 DOI: 10.1038/35097076
Really interesting =)
ReplyDeleteI've been asigned an article about FOXP2 at the University (Master in Bioinformatics) and I can assure your post has been pretty useful for me.
Greetings from Barcelona ;)